ジャーナルクラブ
Molecular basis of the autoregulatory mechanism of motor neuron-related splicing factor 30. (JBC, 2025. Spt.) (志田 達哉)
SPF30とは
SPF30(SMNDC1)は、脊髄性筋萎縮症の原因因子であるSMN(Survival of Motor Neuron)のパラログであり、どちらもTudorドメインと呼ばれる対称性ジメチルアルギニン(sDMA)修飾を受けたタンパク質と特異的に結合する。一例として、U snRNPを形成するSmタンパク質がメチロソーム複合体によりsDMA修飾を受けることにより、SPF30やSMNと結合することが知られている1–3 。
SPF30はU4/U5/U6 tri-snRNPをpre-mRNAのスプライシングを担うスプライソソームへリクルートする因子として同定され、必須のスプライシング因子として知られていた4–6 。しかしその後の研究から、SPF30は必須因子というよりも、特定の遺伝子群や組織特異的なスプライシングで機能する因子である可能性が示唆されている。さらに、SPF30 は RNA分解装置であるRNA エキソソーム複合体(MTR4 など)と相互作用することが示されており、mRNAのみならずncRNAを含む広範なRNA 代謝にも関与する可能性が考えられている7,8 。また、疾患との関連性においては、SPF30 の発現異常と癌や神経疾患の発症との関連性が報告されているものの、その基盤となる病態分子メカニズムは未だ解明されていない10 。
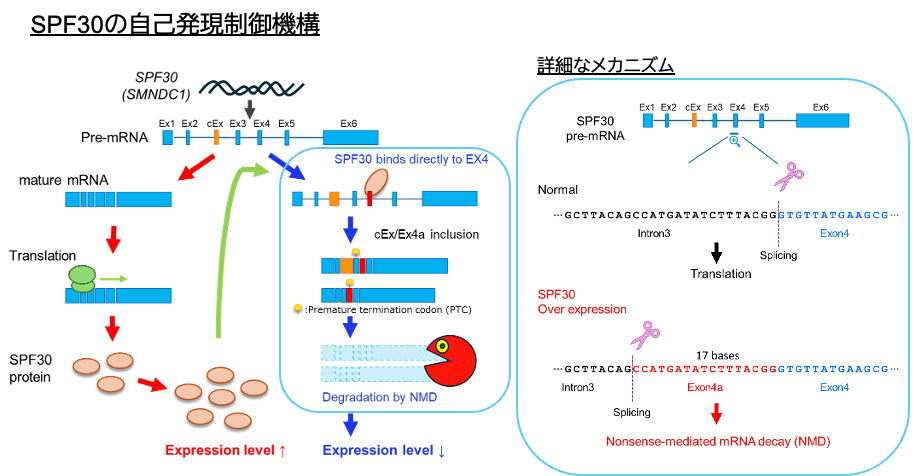
SPF30の自己発現制御機構とは
一部のタンパク質は、過剰発現や機能不全により細胞毒性や代謝異常を引き起こす。このため、細胞内にはそれらの発現量を一定に維持する制御機構が備わっている。この機構には、転写、RNAプロセシング、翻訳、分解など、さまざまな段階での精密なフィードバック制御が機能している。特にスプライシング因子やRNA結合タンパク質の一部は、自身の発現量を直接調節する自己発現制御機構を有しており、細胞内の自身の発現量を一定に保つ負のフィードバック機構において機能している11–13 。
我々はSPF30の自己発現制御機構の分子メカニズムに着目し、その詳細な分子機構を明らかにしてきた。SPF30の自己発現制御機構では、SPF30タンパク質自身がSPF30 pre-mRNAのexon 4部位の5’側領域に結合することで選択的スプライシングを誘導し、既存のexon 4の17塩基上流でスプライシングが起きる。このスプライシング部位の変動により産生されたmRNAは早期終止コドン(PTC)を持ち、ナンセンス介在mRNA分解(nonsense-mediated mRNA decay ; NMD)によって分解される。この選択的スプライシングは、SPF30のタンパク質量が増加した場合に誘導され、タンパク質量が低下すると、通常のexon 4を選択するスプライシングに戻る。その結果として、SPF30 の発現量が一定範囲内に維持される自己発現制御機構が形成されていると考えられる14 。SPF30にはIntron 2領域に存在するカセットエキソンの挿入による別の選択的スプライシング経路を介した自己発現制御機構も存在する15 。興味深いことに、両者の遺伝子配列の保存性を比較した結果、今回我々が新たに同定した自己発現制御経路は、脊椎動物の魚類以降、(別経路は爬虫類以降)で高度に保存されていた。このことは、SPF30は生物種を超えて発現量を厳密に制御すべきタンパク質であることを意味すると考えている(志田 達哉)。